Instructor: Dean Gabriel
a. The frequency of recombination between any
two genes is stable, and is used as the basis of mapping. The percent recombination
is also called a map unit, m.u. or a centiMorgan (cM). Therefore we would
say in the case of the white and frizzled loci that they are 20 cM or 20 m.u.
apart.
b. The maximum distance between two loci that
can be deduced by any single cross is 50 cM.
c. The number of linked groups is identical to
the haploid chromosome number, if the marker density is high enough.
d. Genes are found in linear order on chromosomes
(not branched).
e. The linear order of genes on a chromosome may
be determined from recombination data.
f. The genetic map distance provides some basis
for estimated the physical linear distance.
Work out the following problem for yourself:
Consider 3 genes abc in a haploid parent crossed with a wild type parent. (Fungi are usually used for haploid genetics, and the markers are typically mutations affecting nutritional requirements, such as histidine-requiring or uracil-requiring, etc. These are called auxotrophic mutations, meaning, requiring nutritional supplements that the wild type does not require. [A prototroph is a wild type]):
If abc X +++, and the results are:
abc 67%
+++
a++ 7.5%
+bc
a+c 22.5%
+b+
ab+ 2.5%
+c+
Give the gene order and calculate the distance
between each gene. With this exercise, it can be readily seen that with
enough genes, one could determine that genes are in linear order, and not
branched. Q: What would one expect if chromosomes were branched at
the ends?
Gregor Mendel used peas, Calvin Bridges and Curt Stern used Drosophila, and many subsequent geneticists used fungi to do genetic analyses. The choices were in part driven by temporal concerns: the generation time of peas is about one year, Drosophila is 14-17 days and yeast (a non-filamentous fungus of the class Ascomycetes, is 2 days. Most of the fungi used for genetic analyses are haploid, which makes dominance effects, such as those we saw in the chicken example, above, a non-issue. How do haploid organisms (nuclear state, N) undergo meiosis?
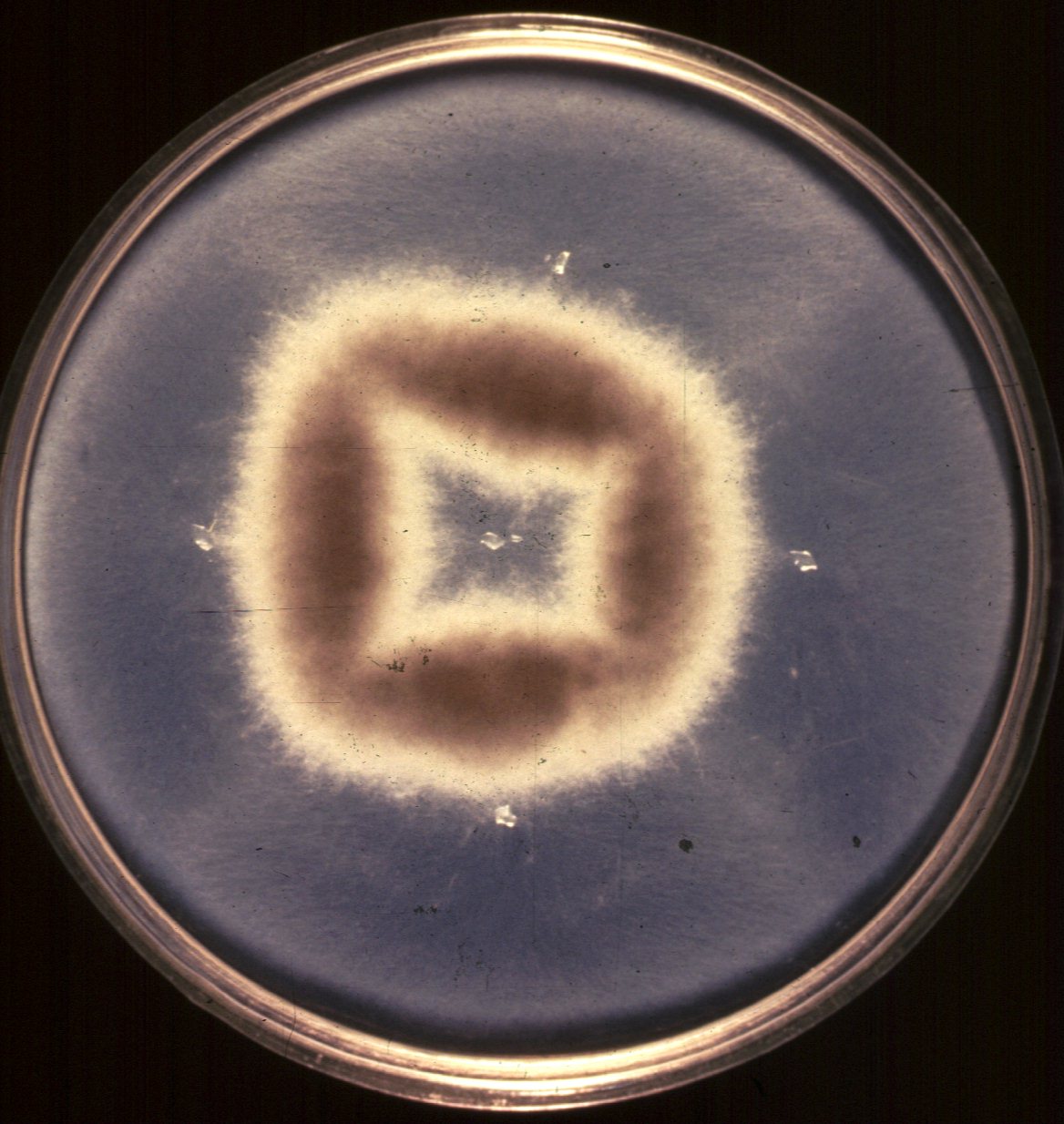
First, the cytoplasms of the two sexually compatible strains (there are no male and female fungi) must fuse. This is called plasmogamy, and creates an N + N state, which is nothing more than two distinct nuclei in a common cellular cytoplasm. (This is similar to fusion of haploid gametes, prior to nuclear fusion). One can force this state rather easily in culture medium by using two strains with different nutritional requirements (auxotrophs). For example, if one strain is auxotrophic for histidine, and the other auxotrophic for methionine, by placing them together on a common medium lacking both methionine and histidine, if their cytoplasms fuse, then the nucleus from the methionine auxotroph provides the enzyme needed to produce histidine, while the nucleus from the histidine auxotroph provides the enzyme needed to produce methionine. The two nuclei are said to complement each other, and the strains now grow very well on minimal medium lacking methionine and histidine.
For example, here is a standard complementation test . photo of a petri dish containing two different auxotrophs complementing each other. The first auxotroph was placed in the middle of the dish, and grew out of the small agar chunk that is visible, and the other auxotroph was placed in four sections surrounding the first, and grew from the four other agar chunks. Notice how the growth of the common cytoplasm (N + N) is luxurient, while the growth around the agar chunks is barely visible.
If the N + N cytoplasm is now placed on meiosis induction media, karyogamy (fusion of the two nuclei) occurs, resulting in a true 2N state. In some fungi, this state is stable, while in others, meiosis usually occurs immediately, resulting in haploidization. If the fungus is an ascomycete, the cells that result from meiosis are called ascospores, and these are usually all readily analyzed. By contrast, one cannot readily analyze each of the gametes of most diploid organisms (try to do this with pollen, sperm or eggs).
Finally, in many ascomycetes, not only are all the products of meiosis available for genetic analysis, but some of them are available in the order in which reductional and equational division took place.
{kind=link}