Instructor: Dean Gabriel
DNA replication -->Pre-leptotene -->Leptotene -->Zyogtene -->Pachytene -->Diplotene -->Metaphase I --> etc.
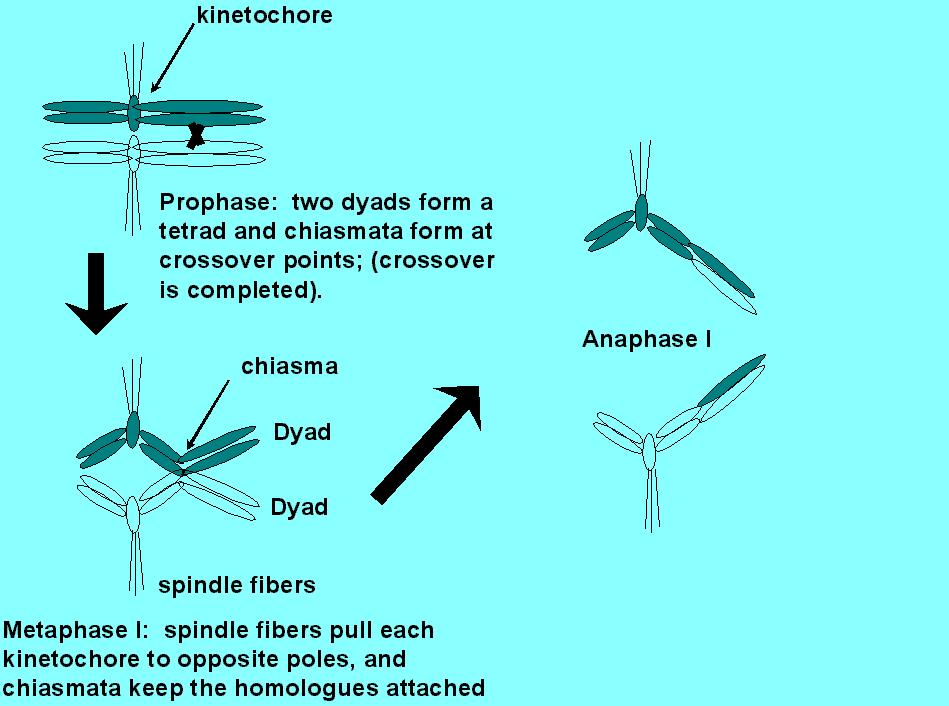
After replication or S phase, each chromatid has a "
sister" chromatid, which together form a dyad. The
sister chromatids of the dyad stay attached along their entire lengths by
a single axis (see below) and by a single shared kinetochore.
A kinetochore is a darkly staining fibrous object that appears to form at
the centromere (and may include a part of the centromere) and directly
attaches the dyad to the microtubules, which comprise the
spindle fibers. The microtubules attach each dyad to opposite
poles of the cell. Although in mitosis each sister chromatid appears
to form its own distinct, darkly staining "axial element", in meiosis,
the sister chromatids of a dyad appear to form a single conjoined sister/sister
axis. This conjoined axis also keeps the sister chromatids attached
to each other during meiosis I. Together the two dyads form a tetrad
, which is also known historically as a "bivalent", principally because
the conjoined sister/sister axes make the tetrad appear bivalent, rather
than the actual tetravalent.
Crossing over occurs in prophase. Dyads line up, side-by-side, in a process called homologue pairing. (Refer Kleckner, 1996). There are multiple, but well spaced, physically observable pairing associations made along the lengths of interacting homologues (from 3-4 to 60-80, depending on the organism), but only a subset of these will result in crossovers . Each tetrad (a.k.a. "bivalent") will most likely experience only one or two crossovers. However, the probability that the tetrad will experience no crossovers is less than 0.1%!
To see a nice set of movies detailing meiosis and crossing over created at the University of California, Santa Barbara, first click on Prophase , then click on Meiosis . Note particularly the details: in the Prophase movie, you see recombination without crossing over, and in the Meiosis movie, you see recombination resulting from crossing over. (To view these movies, you need "QuickTime" Viewer, freely downloadable from: http://www.apple.com/quicktime/ )
Following prophase, crossing over is complete, but not physically obvious. In metaphase I, the regions that were involved in crossing over are the last to separate. These regions are called chiasmata, and refer to the physical attachment regions that are observed in metaphase I. Notice that crossing over is a reciprocal swap of chromatid fragments. Is it likely to be precise? To answer this question, we have to resort to genetic data, and the best genetic data was obtained definitively by analysis of tetrads.
In many organisms, including many fungi and some algae, all the products of a single meiotic event may be recovered and analyzed. That is, the four physical chromatids that were involved in meiosis separate and can be recovered as a group. One group of fungi in particular, the Ascomycetes, has provided a wealth of information about crossing over that was deduced by recombinational analyses of tetrads. In fact, some Ascomycetes are so obliging that they provide each separate chromatid in a spore that is arranged in order of the meiotic divisions! These are called "ordered" tetrads:
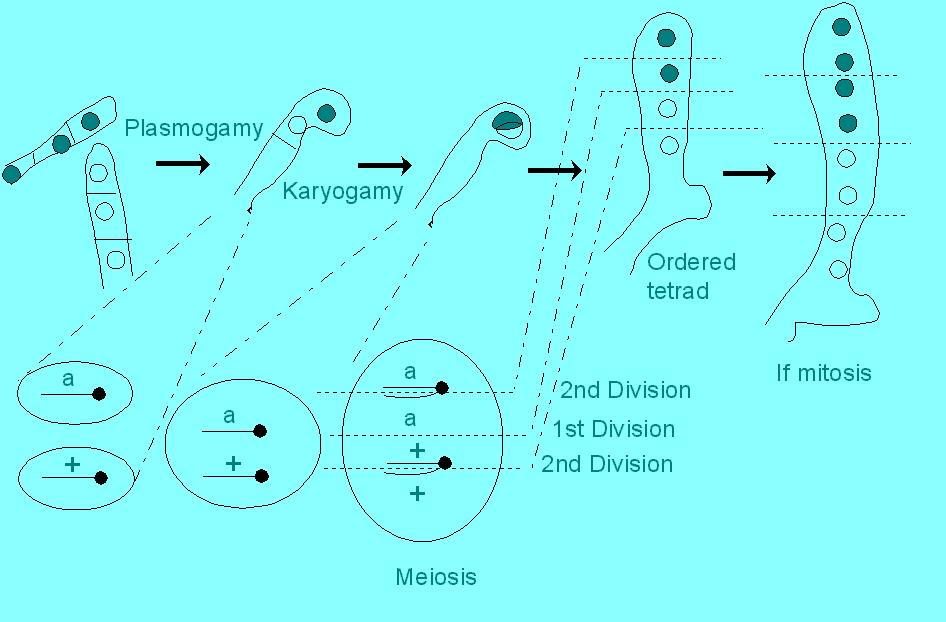
Notice in the Figure above the pattern of dark (phenotype encoded by gene "a") vs. light (wild type) spores. This is a "first division" segregation pattern, and refers to the order of dark vs. light. This depends on the actual orientation of the poles that the dyads are pulled towards in the first meiotic division. If the cell were to have turned upside down, there would be two dark spores down and two light spores up. In the second meiotic division, both sister chromatids that are about to separate at the centromere have either "a" or "+" encoded on them, and so there will be a cluster of two dark and two light. In some fungi with ordered tetrads, a mitotic cell division immediately follows meiosis, and these are also ordered, resulting in a cluster of four dark and four light. Note that it is the kinetochore pulling the centromere that determines the pattern. Therefore, the pattern is a phenotype and the centromere is a "marker" that determines the pattern.
What happens if there is a crossover between the indicated gene "a" and the centromere?
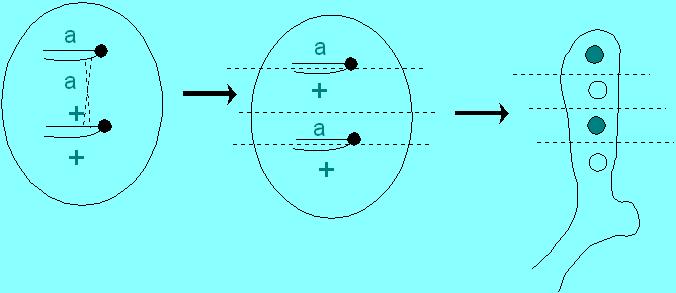
Note that crossing over between the gene and the centromere results in a pattern that is set by the second meitic division; hence it is called a second division segregation pattern.
It can be: a + a + or + a + a or + a a + or a + + a.
Now a question: does crossing over in meiosis occur prior to chromosome replication? Why?
The answer is, NO, and the reason is that if it did, one would only observe first division segregation patterns. The fact that one observes second division patterns proves the principle that crossing over must occur at the four chromatid (tetrad) stage of meiosis.
Now a problem: Consider an analysis of 100 tetrads of the sort illustrated above. Given that 90 have 1st division patterns, and 10 have second division patterns, can you calculate the distance between the "a" gene and the centromere?
One way to answer this is to treat the centromere as a marker gene. Out of 100 tetrads, you will get 400 individuals (4 products of meiosis/ tetrad) or 800 if you have a mitotic division after meiosis, but let's say we don't, so there are 400 individuals total. If 10 have second division patterns, it means that 20 individuals are recombinant between gene "a" and the centromere (remember the second division pattern results from a single crossover involving only two of the four chromatids, and therefore only two of the four resulting individuals will be recombinant). Therefore, 20/400 = 5% recombination.
One can shorten the logic to simply state that the Frequency of 2nd Division patterns, divided by 2, equals the map distance from a given locus examined to the centromere.
Most fungi that produce all meiotic products in a single group produced unordered products, or unordered tetrads. Unordered tetrads are most useful for rapidly determining if two markers are linked.
Consider a two factor cross: a b X + +
If the two genes are unlinked, one expects 0.25 a b, 0.25 a +, 0.25
+ b and 0.25 + + by normal recombinational.analysis.
However, it takes a much larger number of progeny from random sampling of
progeny to obtain linkage data than it does from an examination of tetrads,
whether unordered or ordered. Here's why:
In the cross a b X ++, there are only 3 possible types of tetrads: parental ditype, nonparental ditype, and tetratype:
ab ab
a+
PD = Parental Ditype
ab +b
a+
TT = TetraType
++ a+
+b
NPD = NonParental Ditype
++
++ +b
PD TT
NPD
Note that PDs result from no crossing over, that TTs result from a single cross over, and NPDs result from two crossovers that involve all four chromatids (a.k.a. a four "strand" double CO, where "strand" does NOT refer to a Watson/Crick strand, but rather to an entire chromatid).
To compute recombination frequency between A & B, a reasonable estimation is to add 1/2 of the TT class (of which two of the four progeny are not recombinant) to the NPD class (a four chromatid double CO) and divide by the total:
recomb. frequency between A & B = 1/2TT + NPD
total tetrads
However, this method of calculation underestimates the double crossovers between A & B, and therefore the recombination
between A & B, since it includes only
those double crossovers between A & B that involves all four chromatids. There are also double crossovers between A & B
that involve only two interacting chromatids (in the older literature, "2-strand doubles") that are not recombinant, and double crossovers between A & B that
involve just three chromatids (ie., "3-strand doubles") and are recombinant but only counted once in the above formula, in addition to the "4-strand doubles",
which show up as NPDs. The ratio of 2 chromatid doubles: 3 chromatid doubles: 4 chromatid doubles is 1:2:1. The 2 chromatid doubles are not recombinant and
so are not counted. Since the 3 chromatid doubles are, in fact, double recombinants
but are only counted once in the previous formula as tetratypes, and since these should be present at a 2:1 ratio over the
four chromatid double recombinants (revealed as NPDs), a corrected calculation of recombination distance between A & B, known as the
"Perkins formula" (Perkins D. 1949. Genetics 34:607-626; refer Appendix, p621-622)
triples the NPDs:
recomb. frequency between A & B = 1/2TT + 3NPD
total tetrads
If PDs = NPDs, then the genes are unlinked. This may be concluded
from just a few tetrads. If the genes are linked, the PDs will be
greater than the TTs, which in turn will be greater than the NPDs.
The finding of just a few NPDs out of the first dozen or so tetrads indicates
lack of linkage.