Instructor: Dean Gabriel
Before we can see this violation of independent segregation and how powerful tetrad analysis was in discovering it, we will consider the first question. We know that crossing over occurs at the four chromatid stage (because of what? Second division segregation patterns), but are all of the chromatids involved in crossing over, or only two of them? If all four chromatids are involved in crossing over, are they equally involved, or is there chromatid preference? (this is called "strand" preference in the literature, but "strand" refered to chromatids, not Watson/Crick strands).
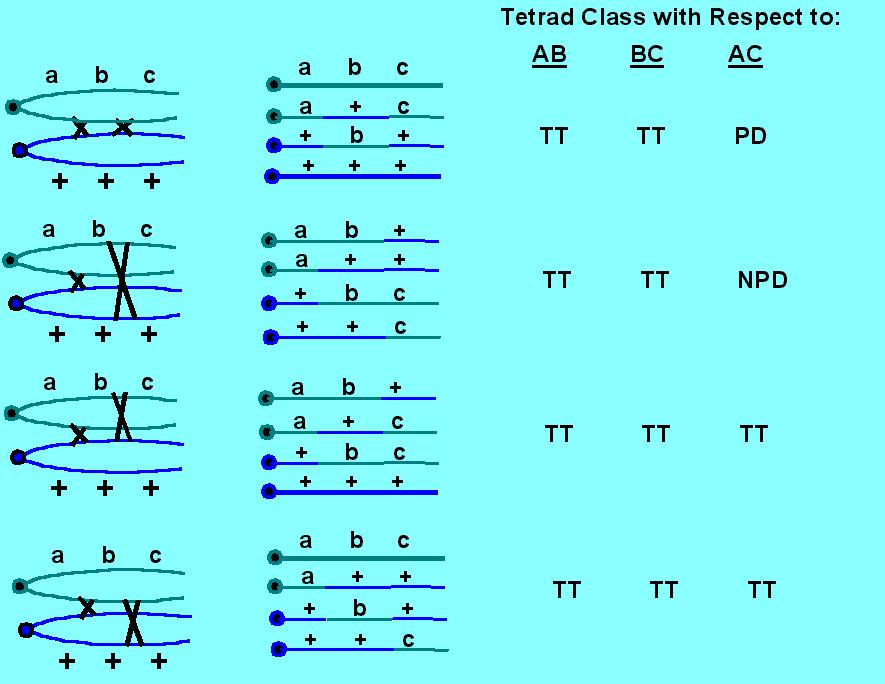
These questions can be attacked by using a three factor cross: abc X +++ , where a, b, & c are linked (genetically, such that PD >> NPD). This allows detection of double cross overs. In the diagram below, consider your expectations given that one crossover (the first one on your left) is a given. That is, the first crossover between chromatids 2 & 3 is fixed. What are your expectations with regard to the second crossover? Here are all the possibilities:
First of all, note that you have to have a 3 factor cross to detect double crossovers, and the experiment can only detect double crossovers in the regions between the markers. Second, note the expectations: one expects double crossovers involving only 2, only 3 or all 4 strands to occur at a frequency of 1:2:1.
Third, note that if you have no crossovers, you get PD (Parental Ditype) for all tetrad classes. If you have one crossover, you get PD for the AB or the BC classes. If you have two crossovers, you get TT (TetraType) for both the AB & BC classes. Knowing that you have two crossovers, one can readily see that the AC class distinguishes among 2, 3 & 4 strand double crossovers. All 2 strand doubles are PD, all 3 strand doubles are TT and all 4 strand doubles are NPD (NonParental Ditype).
The results from several different fungi are that there is a slight, but real, increase in 2 strand doubles over expectations. This allows us to conclude that there is a slight strand preference for crossing over; that is, any two strands that engage in a single crossover are slightly more likely to be involved in a second crossover than any other strands. However, as a rule, we can conclude that all four chromatids are likely to be equally involved in crossing over.
Perhaps the most powerful application of tetrad analyses came as a surprise, with the discovery that at a fairly high frequency, one does not always see independent segregation per Mendel's first law. Sometimes, two alleles go into meiosis, and only one comes out. Sort of like the story of the man and the bear. The man was cold and the bear was hungry and they both ran into the same cave for shelter. A negotiation ensued. At the end of the negotiation, the man exited the cave. It was a win-win situation: the man was no longer cold and the bear was no longer hungry. But I digress. The main point is that unless one can see all four products of meiosis, the occasional disappearance of one of the alleles of a cross would go completely unnoticed in randomly obtained progeny data.
Now we are ready to consider something that only tetrad analyses could (and did) reveal, to the delight and consternation of many: the fact that unequal representation of alleles was sometimes observed after meiosis: That is, people found that very often they would recover 3 a : 1 + or 3 +: 1 a in a cross. Obviously, one wouldn't pick this sort of thing out by looking at progeny in a cross involving Drosophila or chickens.
This phenomenon was called "gene conversion", and is strictly defined as an unequal representation of alleles after meiosis (or mitosis, but we will cover that later). When meiotic gene conversion was examined in some detail, it was found that those fungi that produced a mitotic division immediately following meiosis (such that 8 ascospores, rather than 4, were recovered) provided even more information with regard to gene conversion. Let's consider our 3 factor cross, abc X +++. What were found, occasionally, were tetrads that looked like this :
a b c
Note the 5+ : 3b class.
a b c
+ + c
+ + c
a + +
a b +
+ + +
+ + +
Also found were:
a b c
Note the 6+ : 2b class.
a b c
+ + c
+ + c
a + +
a + +
+ + +
+ + +
Some experimental facts regarding gene conversion:
1. At stable frequencies, characteristic for a locus, alleles may be interconverted.
2. In every locus examined in fungi, gene conversion was observed.
3. Frequencies of gene conversion in meiosis vary from locus to locus, but may be as
high as 1% of the total tetrads examined.
4. One does not find 7:1 or 8:0 conversions, indicating that only two chromatids
are involved in gene conversion events.
5. Gene conversion is accompanied by crossover of outside markers
only 50% of the time!
About this time, people knew:
1) genes were on chromosomes
2) genes were units of heredity
3) crossing over of chromosomes was necessary to obtain independent assortment
4) chromosomes were comprised of DNA & protein.
And then, Watson & Crick published (1953) the structure of DNA. The structure could have been boring. But it was anything but boring. It was already known that DNA was comprised of four nucleotides, but it was the revelation that structurally it was comprised of two intertwined strands of complementary DNA that was startling. This fact immediately suggested that one strand served as the template from which the other could be copied. And this provided a chemical basis for recording, preserving, and recalling hereditary information.
Now people became serious about models that could explain all the "hereditary units" (aka "genetic") data they had been recording.
The first model proposed was partly true: Breakage-Reunion. It was a naive model that didn't help to explain much, but it simply stated that chromosome fragments broke off and reattached. It was dismissed because chromosomes generally have a stable structure. As we shall see later, breakage-reunion does actually occur, and is an essential part of the best model to date.
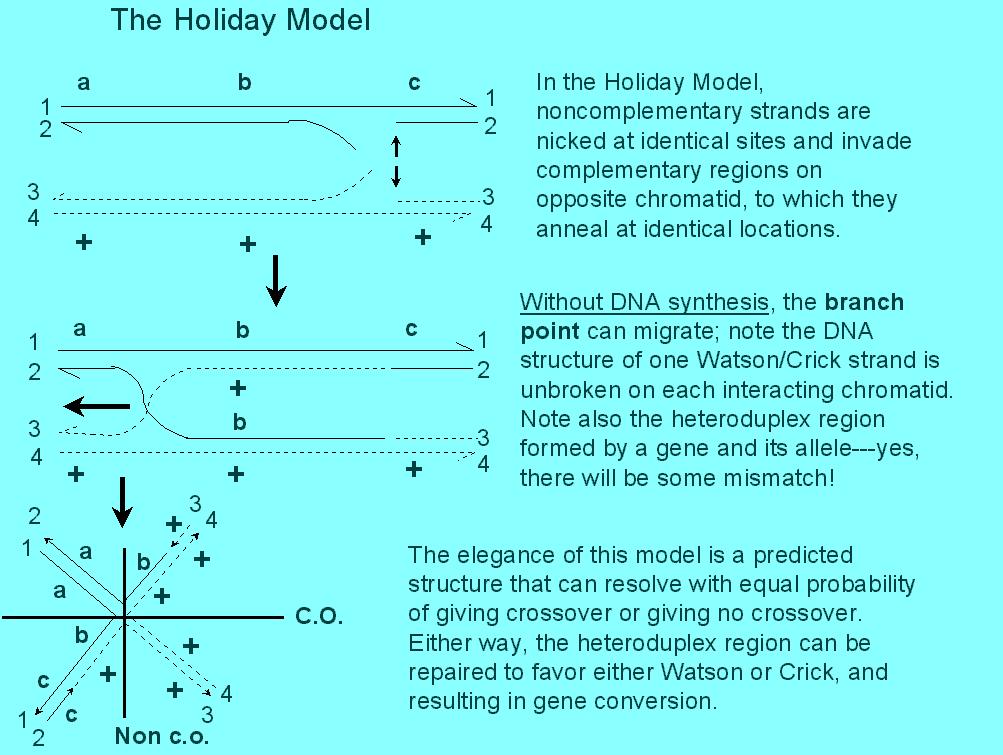
The Holliday Model was the first serious & credible model that explained most of the data. Furthermore, it made certain testable predictions, which is a sign of a good model.
Make sure you can write out the expected products (test yourself):
If crossover: a b +
and + b c
a + +
+ + c
If non crossover: a b c
and + b +
a + c
+ + +
Note that the Holliday model produces two regions of heteroduplex DNA
that are symmetrical. Heteroduplex DNA is that which carries
the DNA coding sequence on one strand, and an allele, with a slightly different
sequence (say, a point mutation, or frame shift, or addition, or deletion)
on the other strand. This would result in a mismatch of base
pairing at one or more points. There should be a mechanism for correcting
or repairing mismatches. In the Holliday model, mismatch repair on one
chromatid would result in 5:3 tetrads, while 6:2 tetrads would result from
two such corrections. Since the frequency of tetrads with 6:2 > 5:3
>>>ab4:4s, Holliday proposed that repair was very efficient.