I. Mendel's laws (published 1866, republished 1900)
A. Offspring looked like parents (phenotypic variation
is inherited), but are not a blend (you get boys or girls, but not (usually)
in between.
B. Characteristics that are well-defined tended to be either/or,
like gender: you get wrinkled or round peas, not semi-wrinkled, you
get yellow or green peas, etc.
C. Mendel defined these either/or units of heredity as
"genes".
D. Working with self-pollinating pea plants, Mendel found
that if the trait bred "true" (eg., the offspring were all like the parents
in that trait), and if variation in the trait was either/or, then genes
had to occur in pairs within each parent, who could pass along one or the
other of the two genes (we would say, alleles) to offspring in the gametes.
E. Try to work it out with the hypothesis that genes don't
occur in pairs. What if they occur singly? What if they
occur in triplets? Quadruplets? (If Mendel had worked with
tetraploid potato, what result would he have seen?
F. The genes that occurred in pairs segregated independently
(Mendel's 1st law). A corollary is that each parent contributes equally
to offspring. (This actually violated the philosophy/theology of
some, who thought that males planted seeds in fertile soil.)
F1 (first filial generation)
F2 (second filial generation; the result of selfing)
RR (round) X rr (wrinkled) ---> Rr (round)
----> 1 RR : 1 Rr : 1 rR : 1 rr
Note 3:1 (R-: rr) ratio in the second filial generation (F2)
E. What about other genes? Do parental types tend to remain together? (If I cross round yellow peas with wrinkled, green peas, does roundness stay correlated with yellow?) No, actually, different traits tend to assort independently (Mendel's 2nd law).
F1
RRYY (round, yellow) X rryy (wrinkled, green) --->
RrYy
(round, yellow)
F2
---> 9 R-Y- : 3 R-yy : 3 rrY- : 1 rryy
(Work out a table of gametes: RY
Ry rY ry
RY
Ry
rY
ry
How do we know that genes are on chromosomes?
F. In 1903, W.S. Sutton, a nuclear cytologist, heard Mendel's theory of genes occurring in pairs and segregating independently, and pointed out that chromosomes do, too.
G. In 1905, EB Wilson and Nettie Stevens demonstrated that gender as a trait was associated with a specific chromosome in insects.
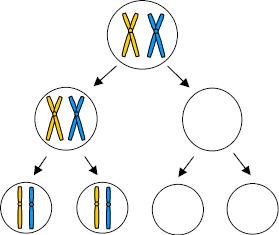
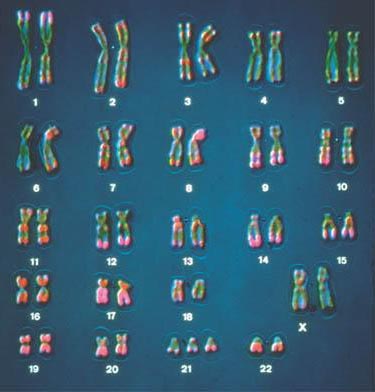
H. In 1916 Calvin Bridges proved that genes were on chromosomes using a combination of cytology and genetics. He relied on the phenomenon of primary nondisjunction [Failure of chromosomes to separate at metaphase I (reductional division)]. Illustrated below left is a diagram of primary nondisjunction in meiosis. Illustrated below right is the human consequence of primary nondisjunction, resulting in trisomy 21, the most typical cause of Down's Syndrome. (Note that each of the condensed, compact chromosomes that are readily visible are metaphase cells deliberately arrested during mitotic division following the S phase of chromatid replication, resulting in a pair of sister chromatids attached to each other at the centromere. Each of these pairs of sister chromatids is called a "dyad", and although there are two different copies of each chromosome (2N), the dyads represent duplicated copies, sometimes referred to as a "4C" state.
In his first experiments, Bridges used white eyes, a recessive trait, and performed a cross with red eyed males.
Make a table of gametes and predict the results:
w+ Y
w w+w w Y 49.95%
w w+w w Y 49.95%
w/w dead w/w Y 0.05%
Ø w+ dead 0.05%
Note that we expect all females to be red eyed, and all males to be white eyed. The exceptions are white eyed females and red eyed males. Bridges demonstrated that all of his genetic exceptions also had the predicted chromosomal abnormalities. (The white eyed females had a Y chromosome; the red eyed males had a single X and no Y chromosome). Some had argued that the exceptions were mutational and that the genes were cytological, not nuclear. Bridges showed that all exceptions had chromosomes expected of primary nondisjunction. Some now argued that a mutation caused both the chromosomal conditions and the genetic exceptions. Bridges countered by predicting that any sex-linked trait would show the same behavior, and set up a similar experiment using four different genes:
x, y, z are recessive; Bar eye (B) is dominant. He started with xyz nonBar males, crossed to wild type females, selected the female offspring, and crossed them with Bar eyed males: (Make a table of gametes, and be sure to include what you would expect if you had primary nondisjunction. In the table below, note that the first two rows are expectations if reductional and equational division work normally in meiosis; the last two rows are expectations after primary nondisjunction.)
Parents: xyz+ X
+++B
++++ Y
+++B Y
++++ +++B
++++ Y
++++
xyz+ +++B
xyz+ Y
xyz+
++++ dead
++++ Y
xyz+
xyz+
Ø +++B
dead
Note that we expect all wild type, Bar eyed females, and all non- Bar eyed males (50% being triple mutant and 50% wild type for xyz) Exceptions are easily identified as non Bar females and Bar eyed males. Only genetic exceptions showed an unusual chromosome condition, and 100% of the Bar eyed males were wild type for x,y and z. Also note that only the exceptional females, which exhibited primary nondisjunctional, also showed no segregation of genes. Segregation of four independent genes failed in the exceptions only. This could only happen if genes were on the chromosomes.
The proof that a physical exchange of chromosome pieces or a crossover was needed for recombination of genes on a given chromosome was provided by Curt Stern in 1931. The proof involved use of two aberrant looking X chromosomes that were created by irradiating Drosophila. The female parent carried both aberrant chromosomes, one of which carried the recessive carnation (car) colored eyes mutation and also Bar, the dominant Bar eyed mutation. The male parent had a cytologically normal X chromosome marked with car, but not Bar. The gametes from the cross may be pictured:
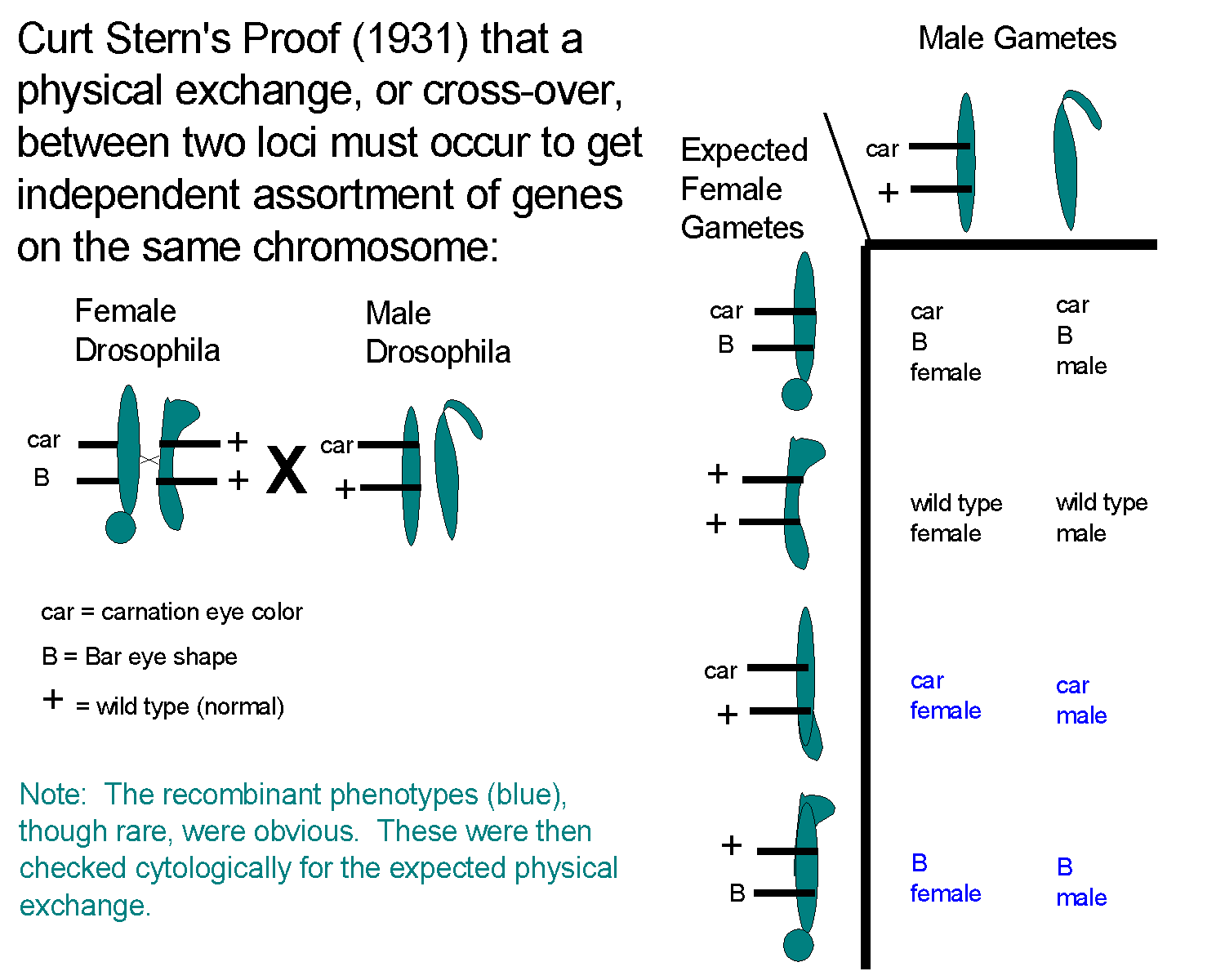
ww, FF
red, frizzled (true breeding, so we know are
homozyous; gametes are wF only)
X
WW, ff
white, nonfrizzled (also true breeding, gametes
are Wf only)
yields F1: WwFf (white frizzled)
We can't self-cross these, as Mendel could with peas, so we have to resort to a "test cross" (this means use a double recessive individual as a tester, to eliminate dominance effects):
Ww, Ff
X
ww, ff (red, nonfrizzled tester)
yields:
Possible Expected Actual
WwFf 25% 18
wwFf 25% 63
Wwff 25% 62
wwff 25% 13
156
Ignoring the contribution of the tester's gametes,
note the tendency of the original parental gamete combinations wF &
Wf to remain together. This is the definition of linkage:
the tendency of parental gene combinations to be inhereted as a unit. Note,
too, that the new combinations of two genes not originally seen
in the parents is 20% of the total. This is the definition of recombination:
new combinations of two genes not found in the parental gametes. (What
is the expected recombination frequency? Can recombination frequencies
between any two genes ever be higher than 50%?)